'
Naar een herziene aardgeschiedenis (geologie)?
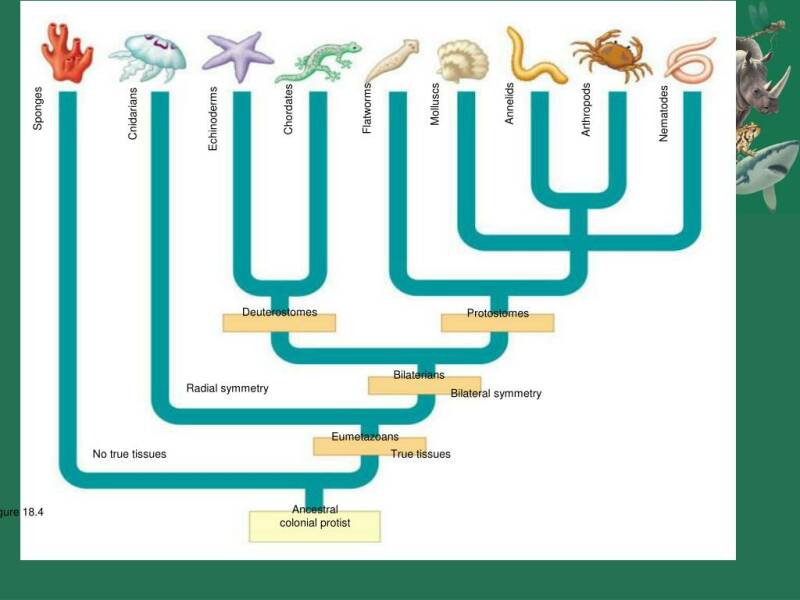
In het boek van Jens Boeningk, Sabine Wodniok en Edvard Glücksman Biodiversity and Earth History wordt een prachtig overzicht geboden van alle geologische perioden en de bijbehorende diverse levensvormen. Bij de indeling van de organismen wordt uitgegaan van het cladistische systeem.
De drie gezamenlijke criteria hiervoor zijn:
- de structuren van DNA en RNA
- stofwisselingsprocessen
- morfologische overeenkomsten en verschillen.
Bij de bespreking en beoordeling daarvan ga ik eerst de complete evolutionaire indeling langs. Het einde van de periode Krijt wordt gemarkeerd wordt door een geweldige catastrofe waarbij er een einde kwam aan het Dinosauriertijdperk. Door de inslag van, zo meent men op goede gronden, een enorme meteoriet. Tijdens de Krijtperiode waren alle groepen van het dieren- en plantenrijk reeds aanwezig.
De supergroepen.
(de structuur van de cellen heb ik getekend naar het boek Biodiversity and Earth History).
1 Unikonta.
Hiertoe worden gerekend de Ophisthokonta (de flagel achterwaarts gericht), de Amoebozoa (amoeben en slijmzwammen met schijnvoetjes; ze hebben wel of geen flagel) en de Apusozoa. Alle "dieren" zijn unikont.
De fylogenetische relatie binnen de diverse groepen van Unikonta is zeer onzeker. En: de indeling van alle organismen op grond van het aantal flagellen is erg onbetrouwbaar.
1.1 Holozoa.
De Holozoa omvatten zeer heterogene groepen zoals 1-celligen, sponzen, insecten, wormen, schimmels, vissen, zoogdieren en mensen.
De fylogenetische ontwikkelingslijnen binnen de Holozoa blijven nog steeds onduidelijk!
Vaak zijn er exoskeletten ontwikkeld; al ver voor het Cambrium. Tijdens de zgn. Cambrische explosie.
a. Choanomonada.
Eén- of meercellige organismen met 1 flagel. De flagel heeft 2 tubuli in het midden en 9 tubuli daar omheen. De flagel is omgeven met 20 à 40 microvilli die actine-filamenten (vgl. onze spieren) bezitten. Ontstonden de Metazoa als taxon vanuit de Choanomonada of is er sprake van een monophyletische groep of van een paraphyletisch groep?
Vaak is een kinetosoom aan de basis van de flagel aanwezig. Dus een gereduceerde flagel?
b. Porifera (sponzen).
Porifera hebben geen zenuwcellen of spierweefsel. Geen darmkanaal of voortplantingsklieren. Kraagcellen (met 20 à 40 filipodia) zorgen voor voedselopname. Kalk- of kiezelnaalden worden gevormd in de buitenste cellaag.
d. Cnidaria (koralen, poliepen, kwallen en zeeanemonen).
Ze zijn radiair symmetrisch en bezitten netelcellen op hun tentakels Ze hebben een netvormig zenuwstelsel met een zenuwring rond de mondopening. Geen spiercellen.
Fossiele koralen bestaan vanaf het Ordovicium.
e. Ctenophora (ribkwallen).
Ze zijn niet-symmetrisch en hebben geen netelcellen. Ze vangen hun prooi m.b.v. colloblasten. Deze bevatten een opgerolde spiraal in de cel die naar buiten kan schieten en zich hecht aan een prooi.
Ze bezitten spierweefsel en hun ontwikkeling vertoont overeenkomst met de tripoblasten.
Ze bezitten lijsten van trilharen voor de voortbeweging.
Geen fossielen en de phylogenetische plaats is onduidelijk.
1.2 Protostomia
De Protostomia worden beschouwd als een zustergroep van de Deuterostomia.
Bij de Protostomia ontstaat de mond uit de blastopore (de opening van de blastula).
Bij de Deuterostomia ontstaat de anus uit de blastopore.
Fylogenetische relaties binnen de Protostomia blijven onopgelost!
Anneliden (Spiralia) en Arthropoda vertonen overeenkomsten betreffende de gesegmenteerde lichaamsbouw, een touwladderzenuwstelsel en lengtespieren. Zijn ze ook vergelijkbare voor wat betreft hun extremiteiten??
Zijn de monddelen en gelede poten van de insecten dan homoloog met de parapodia van de wormen?
Moleculaire studies weerspreken de verwantschap.
a. Ecdysozoa.
Deze dieren bezitten een driedelig chitine-skelet. Ze hebben een door hormonen bestuurde ontwikkeling via vervellingen. Aangepaste flagellen hebben een sensorische functie en liggen in de cuticula. Ze hebben geen trochophoralarve.
Deze groep omvat rondwormen, geleedpotigen (insecten, spinnen, kreeften), duizendpoten e.a.
De spoelwormen of rondwormen (25000 soorten beschreven maar omvat misschien wel 10.000.000 soorten) hebben 4 zenuwstrengen in de lengte (dorsaal, ventraal en lateraal) en een ring rond de pharynx.
Ze zijn niet gesegmenteerd. Ze hebben wel mannelijke of vrouwelijke voortplantingsorganen.
Het lijkt me onverantwoord om rondwormen in één groep te plaatsen samen met de Arthropoda.
Tot de Tardigrada behoort o.a. het Beerdiertje. Duidelijk gesegmenteerd.
Het diertje kan leven onder de meest denkbare extreme omstandigheden: van +149 graden C tot -272 graden C. Ze verdragen Uv-licht en gammastraling. En dagenlang verblijf in de ruimte.
De fylogenetische plaatsbepaling van de Tardigrada is welhaast onmogelijk vanwege tegenstrijdige resultaten van morfologische en moleculaire gegevens.
De Arthropoda zijn gesegmenteerd met een 2-delig (spinnen en krabben) of een 3-delig lichaam (insecten) of ze bestaan uit vele segmenten (Myriapoda (miljoenpoten en duizendpoten).
Tot de Arthropoda behoren eveneens de beroemde fossiele Trilobieten. Levend tijdens het allereerste begin van de wereldgeschiedenis.
b. Spiralia.
Zo genoemd omdat de eerste delingen van de eicel een karakteristieke (spiraalvormige) klieving vertonen. Daarbij worden 4 macromeren en 4 micromeren gevormd. Daarop komen echter ook uitzonderingen voor!
Tot de Spiralia behoren de Platwormen (zonder coeloom) en Rotifera (met ronddraaiende trilharen).
Tot de Spiralia worden gerekend:
1 een zeer diverse groep van kleine wormachtige dieren (Gastrotricha etc.).
Zonder coeloom.
2 Brachiopoda: nu nog 200 soorten. Veelal levend in diepe zeeën.
Bezitten een lofofoor als krans rond de mond om voedsel te vangen.
Bestaan vanaf Precambrium en overheersen tijdens het
Paleozoïcum. Ze bezitten een onder- en bovenschelp i.t.t. de
mollusken die een rechter- en linkerschelp hebben en zich
voeden m.b.v hun kieuwen met daarop trilharen.
3 Nemertea: soms zeer lange wormachtige dieren (tot 35 m.)
met een proboscis. Voortbeweging door ciliën. Fossielen
vanaf Cambrium.
4 Annelida: de welbekende regenwormen, zeepieren en Nereis.
5 Mollusca: met of zonder schelp: de slakken, 2-kleppigen, inktvissen en
chitons.
De Inktvissen komen voor vanaf het Cambrium. Dit geldt eveneens
voor de Anneliden, Mollusken en Slakken.
Conclusie van de schrijvers: de onderlinge relaties binnen de polyphyletische groep van Spiralia zijn volledig onzeker. Moleculaire studies plaatsen b.v. de Brachiozoa naast de Annelida.
De Mollusca vormen een geheel van onherleidbare groepen. De schelpdieren (2-kleppigen, slakken en inktvissen) zouden een monophyletische groep zijn maar de diverse schelptypen zijn totaal verschillend.
1.3 Deuterostomia.
De eerste opening van de blastula ontwikkelt zich tot anus.
Hiertoe behoren de Chordata (waartoe ook de vissen, vogels, zoogdieren en mensen behoren) en de Ambulacralia (waartoe de Echinodermata en de Hemichordata behoren).
a. Hemichordata.
De Hemichordata vormen een wonderlijke groep omdat daar de uitgestorven, oeroude groep van Graptolieten gerekend wordt; levend vanaf het Cambrium. Een heel bijzondere groep is de wormvormige Enteropneusta (eikelwormen). Sommige celgroepen tonen verwantschap (?) met die bij "hogere" dieren die de ontwikkeling van hersenen bevorderen.
Ze hebben kieuwaanleg, hart en bloedvaten evenals een dorsale en ventrale zenuwstreng. Al met al worden ze soms beschouwd als voorouders van gewervelden of als overgang tussen gewervelden en ongewervelden.
Chordata bezitten een dorsale weefselstreng, de Notochord; een zenuwstreng gelegen boven de chorda.
De chorda functioneert als endoskelet en ontstaat uit het mesoderm. Ze komt ook voor in de larven van Urochordata.
b. Echinodermata.
Tot de Echinodermata behoren naast zeesterren, zee-egels enz. de tijdens het Precambrium levende Crinoidea (zeelelies). Sinds 1864 zijn er enkele nu levende soorten van deze laatste ontdekt op de diepe oceaanbodem.
Echinodermata bezitten een ambulacraal systeem. Dit is een schitterend systeem van kanaaltjes door het hele dier, verbonden met de loopvoetjes. Volkomen uniek in het dierenrijk! Niet te herleiden tot enig ander orgaanstelsel!
c. Cephalochordata.
De Cephalochordata worden beschouwd als een belangrijke fossiele groep die nu nog voort bestaat als Amphioxus (lancetvisje). Ze voeden zich door filteren van water dat over de kieuwen stroomt.
Beroemd fossiel: Pikaia.
d. Urochordata = Tunicata.
Alleen in de larven is de chorda en zenuwstreng aanwezig. Ze leefden al tijdens het Cambrium.
Ze zijn een zustergroep van de Craniata. Maar de relatie tussen Cephalochordata, Urochordata en Craniata blijft een groot probleem!
1.4 Craniata.
Hiertoe behoren de Myxini (slijmprikken en prikken of lampreien). Dit zijn Agnatha.
Overigens behoren tot de Craniata (schedeldieren) alle Vertebrata, inclusief de mens.
a. Gnatostomata.
De kaken bestaan uit beweegbare skeletdelen. Kaken zouden ontstaan zijn uit de 3e en 4e
kieuwbogen.
De Placodermi waren bepantserde bodemvissen die leefden tijdens Cambrium en vooral het Devoon.
De Chondrichtyes zijn de kraakbeenvissen; haaien, roggen etc.. Ze bestaan al vanaf de vroegste tijden: 430 miljoen jr. geleden. Ze vertonen diverse vormen van voortplanting: ovopaar, ovovivvipaar en vivipaar.
De Actinopterygii zijn de echte, straalvinnige vissen met schubben enz.
De Sarcopterygii worden beschouwd als de stamoudsten van de coelacanthen en de longvissen.
Sarcopterygii (lobvinnige vissen) bestonden al vanaf Devoon, 420 miljoen jr. geleden.
Daarmee schijnt de route naar het ontstaan van dieren met longen die lopen op 4 poten aangevangen!
Opm.: Al de bovengenoemde overgangen en gesuggereerde evolutielijnen zouden nog altijd op de één of andere manier bewezen moeten worden vanuit de biologie!
b. Amniota.
Nadat de Lissamphibia waren ontstaan via Lepospondyli of Temnospondyli (levend gedurende het Carboon) kwamen hieruit de welbekende kikkers, padden en salamanders voort, zo meent men. Ze komen al voor vanaf het Devoon. Vanwege ontwikkeling van de eieren in het water is de bevruchting normaliter uitwendig en bezitten de eitjes geen schaal.
Als de voortplanting onafhankelijk van water op het land plaats gaat vinden bij reptielen, vogels en zoogdieren is het voorkomen van een amnion vereist die omgeven is door een leerachtige (bij reptielen) of een kalkachtige schaal (bij vogels)
Andere mogelijkheid: eitjes die zich zonder schaal ontwikkelen in een baarmoeder.
De Archosauromorpha omvatten diverse Sauriërs, krokodillen en vogels.
Tot de Protheria of Monotremata behoren de eierleggende zoogdieren: mierenegels en vogelbekdieren. Ze worden ook wel cloacadieren genoemd. Dus verwant aan reptielen? Maar ze produceren melk!
Alle Theria zijn vivipaar. Hun jongen ontwikkelen zich in het moederlichaam.
Een bijzondere groep naast de Eutheria zijn de Marsupialia (buideldieren). Bij deze dieren verlaat het jong in een zeer vroeg stadium de baarmoeder om zich verder te ontwikkelen in de buidel, samengegroeid met een melktepel. Ze zijn waarschijnlijk ontstaan in de Jura. De eerste fossielen zijn gevonden in het Krijt.
De schildpadden zijn Anapsida. Daarmee behoren ze tot een uitgestorven groep van Sauriërs. Zonder temporale openingen achter het oog. Moleculaire studies plaatst de schildpadden bij de Diapsida waartoe veel andere Sauriërs, slangen, krokodillen en vogels behoren.
Belangrijk discussiepunt binnen de evolutieleer
is warmbloedigheid (homeothermie). Kwam dit al voor bij de Archosauriers en uitgestorven krokodillen? De laatsten hebben een vierdelig hart en een secundair verhemelte.
Opm.: Al de gesuggereerde ontwikkelingslijnen tussen de basisgroepen van de Amniota berusten op wensdenken. Vooral de plaats van de moderne vogels is hoogst merkwaardig. Ook al suggereren gevederde Sauriërs de juistheid ervan.
1.5 Holomycota.
Schimmels hebben geen plastiden, wel mitochondriën. Ze vormen mycelium met 2 kernen.
Schimmels ruimen alle organische stoffen op in de natuur samen met bacteriën.
Een tweede zeer essentiële functie van schimmels is de vorming van mycorrhiza rond de wortels van planten; vooral bij bomen en orchideeën. Vanwege het feit dat de celwanden bestaan uit chitine, cellulose en glucaan.
Basale groepen zoals de Chitrydiomycota zijn 1-cellig en hebben een zweephaar.
Alleen de Chytridiomycota vormen cellen met een flagel (zoosporen en gameten). De sporen van Microsporidia hebben 2 kernen en een binnenin opgerolde flagel die naar buitenschiet en contact maakt met het epitheel van de darmwand van een gastheer. Kern(en) en cytoplasma worden in de gastheercel geschoten. Een volledig uniek verhaal!
De hogere Fungi hebben, denkt men, secundair gereduceerde flagellen.
De Nucleariidae zijn cellen die zich voortbewegen met amoeboïde schijnvoetjes.
De grote subgroep naast de "dieren" vormen de Holomycota (de Fungi).
Want ze zijn 'unikont'.
Chitine verbindt Fungi met de Arthropoda en de basale groepen hebben 1 secundair gereduceerde flagel (unikont). Chitine vertoont grote overeenkomst met cellulose. Eén OH-groep is dan vervangen door een acetamide-groep.
Opm.: ß-chitine is aanwezig in het skelet van pijlinktvissen. Het is daarom zeer de vraag of bezit van chitine reden is om de wieren onder te brengen samen met de 'dieren' in één domein, de Unikonta.
De Amoebozoa hebben geen vaste celvorm; ze vormen pseudopodia (schijnvoetjes). Amoeboïde cellen komen in diverse supergroepen (domeinen) voor.
Ze bezitten geen plastiden. Ze leven wel vaak in symbiose met algen; b.v. koralen en sponzen.
a. Glomeromycota.
De Glomeromycota vormen de mycorrhiza rond plantenwortels. Ze zijn polyfyletisch.
Ze vormen clusters of draden die wortels van planten binnendringen. Ze leven in obligate symbiose met mossen, varens en zaadplanten. Ze zijn dus zo oud als het begin van de aardbolgeschiedenis (vanaf 440 miljoen jr. geleden). Deze schimmels zijn niet soortspecifiek en leven samen met 80% van alle landplanten.
Belangrijk: schimmels bestonden al 900 miljoen jr. geleden. Waarmee leefden ze dan samen? Dus toen ook al chitine?
Misschien met cyanobacteriën? Er bestaat een schimmelsoort Geosiphon die in endosymbiose leeft met de cyanobacterie Nostoc. Let wel: endosymbiose!
b. Basidiomycota.
Ze hebben Celwanden met chitine. En daarom in dezelfde supergroep geplaatst als de Arthropoden.
De roesten of Pucciniales (branden) zijn kwaadaardige plantenparasieten. Ze dringen planten binnen via intercellulaire holtes en vormen haustoria. Ook de Ustilaginales zijn obligatoire plantenparasieten. Ze vormen tumorachtige gallen in b.v. Zea (maïs).
c. Ascomycota.
Celwanden met chitine en glucaan.
Truffels, Penicillinum, Pneumocystis (een longparasiet), meeldauwsoorten, Saccharomyces (bakkersgist) en Clasviceps purpurea (het giftige moederkoren in graan) behoren hiertoe.
d. Amoebozoa.
De Amoebozoa vormen een zustergroep van de Ophistokonta, waartoe alle dieren en schimmels behoren .
De Amoebozoa hebben een flagel (meestal secundair gereduceerd) achterwaarts gericht i.t.t. de Ophistokonta die de flagel voorwaarts hebben gericht.
Amoebozoa hebben een amoeboïde voortbeweging en geen vaste celvorm. Soms met skelet.
Ze bezitten een actine-myosine-celskelet (vgl. onze spieren).
De ontstaansrelaties binnen de heterogene groepen zijn onbekend.
e. Conosa.
Ook een zustergroep van de Ophistokonta.
Een aantal taxa hebben flagellen.
Hiertoe behoren de slijmzwammen. Ze vormen vruchtlichamen vanuit een meerkernig mycelium.
2 Excavata.
De "mond" heeft een specifieke vorm; de zgn. cytosoom. Maar vele groepen missen deze eigenschap of ze is secundair gereduceerd. Een cytosoom komt trouwens ook bij de Ciliata (b.v. pantoffeldiertje) voor.
De flagel is achterwaarts gericht. Groot verschil in aantal flagella: 1, 2, 4 of veel (tot wel 1000).
Sommige groepen zijn amoeboïd qua leefwijze en voortbeweging.
Als ze op anaërobe plaatsen leven zijn de mitochondriën gereduceerd of afwezig. Ze zorgen in het laatste geval nog wel voor de synthese van eiwitten en vetten.
Ze leven grotendeels als commensaal of parasitair.
De Euglenida (groene oogdiertjes) behoren hiertoe. Met of zonder bladgroenkorrels. Met 2 flagellen waarvan er 1 gereduceerd is.
De opslag van reservevoedsel vindt plaats in de vorm van ß 1,3-glucaan.
Dit gebeurt ook in de Haptophyta. Maar die behoren tot de supergroep van Hacrobia.
2.1 Metamonada.
Ze hebben 2 kernen en 2 kinetiden (microstructuren voor de beweging van de flagellen). Per kinetide zijn er 4 flagellen; 3 voorwaarts gericht en 1 achterwaarts.
Ze leven allemaal anaeroob en hebben daarom geen mitochondriën. Ze bezitten wel genen voor de aanleg daarvan. Ze leven veelal als commensalen in insecten. Bij de termieten spelen ze een belangrijke rol bij de vertering van lignine en cellulose. Vele soorten zijn pathogeen: b.v. Trichomonas vaginalis die 170 miljoen mensen per jaar infecteert. De infectie behoort tot de zgn. SOA's (seksueel overdraagbare aandoeningen).
2.2 Discoba.
Sommige groepen met meer of minder flagellen, andere met amoeboïde vormen van voortbeweging. De laatste soms weer met stadia die wel flagellen hebben.
Maar er zijn steeds kleine verschillen met de slijmzwammen van de Amoebozoa.
a Euglenida.
De Euglenida hebben een pellicula. Dit is een stevige structuur van eiwitten onder de celmembraan die de hele cel spiraalsgewijs omgeeft.
De systematische plaats van verschillende taxa is onduidelijk.
Verondersteld wordt dat de Euglenida afstammen van fagotrofe (heterotrofe) organismen die door endocytobiose aan hun plastiden kwamen.
a 1 Kinetoplastea.
Ze hebben een kinetoplast: een DNA-rijke streng door het hele mitochondrium zo lang als de hele cel. De kinetoplast is verbonden met de basis van de flagel.
Er komen zowel vrij levende groepen voor als parasitaire. De vrij levende met 2 flagellen. De parasitaire soort is o.a. Trypanosoma, de beruchte verwekker van slaapziekte, met een ingewikkelde levenscyclus via insecten.
Opm.: De Excavata vormen een zeer heterogenen domein met vele leefwijzen, veel variatie in aantal flagellen en kinetoplasten, amoeboïde voortbeweging, typen mitochondriën en wel/geen plastiden etc. Dus geen afstammingsreeksen te postuleren!
3 Archaeplastida.
Archaeplastida hebben chlorofyl a, zetmeel als reservevoedsel en cellulose als celwandmateriaal. Soms zijn er phycobillisomen aanwezig. Deze phycobillisomen bestaan uit een eiwitcomplex die pigmenten bevatten die geel en groen licht kunnen absorberen
De mitochondriën hebben platte cristae.
De Archaeplastida vormen een homophyletische groep binnen de Cyanobateria omdat de eerste stap in hun evolutie bestond in de endocytobiose van een Cyanobacterie.
Daarna kan er endocytobiose hebben plaats gevonden van een eukaryotische alg.
De Viridiplantae bezitten chlorofyl b in de plastiden. De Streptophyta omvatten zowel een parafyletische groep algen als alle landplanten (Embryophyta).De landplanten bestaan uit de parafyletische groep mossen en de monofyletische groep van paardenstaarten, varens en zaadplanten.
De Glaucocystophyta vertonen overeenkomst met de cyanobacteriën vanwege peptideglycaan tussen de membranen van de plastiden. Dit zou een relict van de celwand van een cyanobacterie zijn. Ze hebben eveneens blaasjes onder de celmembranen. Deze zijn gelijk aan de alveoli van de Alveolata die behoren tot de supergroep van Chromalveolata.
De Glaucocystophyta worden beschouwd als belangrijke overgang vanaf de Cyanobateria omdat ze peptideglycaan bezitten tussen de membranen van de plastiden.
Cyanophora paradoxa wordt beschouwd als een levend fossiel vanwege de blijken van primaire endocytobiose als een 'enkelvoudige gebeurtenis die 1 miljoen jaar geleden plaats greep. Opm.: je zou deze gebeurtenis toch wel graag eens bewezen willen zien!
Rhodophyta hebben een vergelijkbare celstructuur als de Glaucocystophyta maar met enkele bijzonderheden: als enige protist met plastiden bezitten ze fycoerythrine. Het chloroplast-DNA is niet circulair en reservevoedsel wordt opgeslagen als alpha-1,4-glucaan (vgl. amylopectine). Er zijn 3 generaties: gametofyt, carposporofyt en tertrasporofyt.
De spermacellen hebben geen flagel maar bewegen zich amoeboïd. Ze worden gevormd in vele gametangia. Terwijl in 1 vrouwelijk gametangium slechts 1 eicel gevormd wordt.
Hun weefsels bevatten pseudoparenchym en cellulose. Ze hebben zowel chlorofyl a als phycobillisomen en fycoërythrine. Daardoor kunnen ze tot 250 m. diep groeien waar slechts 0.001 % van het daglicht aanwezig is.
De roodwieren bestaan al sinds 1.2 miljard jaar geleden.
De Viridiplantae (de planten) vormen een monophyletische groep. Maar ze is zeer divers in morfologie, celstructuur, levenscyclus, voortplanting en biochemie. Veelcelligheid ontstond vele malen onafhankelijk van elkaar! B.v. Volvox. Volvox heeft 2 celtypen: 1 voor de voortplanting en 2 voor de voortbeweging.
De Chlorophyta bezitten chlorofyl a en b, plastiden met 2 membranen, (gevolg van primaire endocytobiose), cellulosewanden en mitochondriën. Met 17.000 soorten komen ze vrijwel overal voor.
Ze zijn zeer veelvormig: met of zonder flagellen, wel of geen kolonievorming, wel of geen celdifferentiatie, vertakt of onvertakt, draadvormig of bladachtige thalli (b.v. zeesla), wel of geen phycoplast-vorming tijdens de celdeling, wel of zonder schaal, met of zonder endosymbiose (met Fungi als Lichenen). Soms komt veelkernig weefsel voor, zoals bij Ulvophyceae.
Opm.: Opstellen van afstammingsreeksen blijkt onmogelijk.
De Streptophyta omvat diverse groepen parafyletische zoetwater-groene-algen.
De voorlopers; wie waren dat? Wellicht 1-cellige flagellaten met plastiden? Vanuit de Streptophyta ontwikkelden de verschillende vormen van landplanten zich zoals mossen, varens, paardenstaarten en bloemplanten. In de voorbereiding daartoe bezitten ze:
- een celwand van cellulose
- veelcellige bouw
- vorming van een phragmoplast tijdens celdeling
- bezit van plasmodesmata
- apicale groei
- driedimensionaal weefsel
- asymmetrische celdeling
- celdifferentiatie.
- biochemisch en fysiologische overeenkomsten zoals de vorming van GAPDH
(glyceraldehyde-3-fosfaat-dehydrogenase) onder besturing door het
enzym thioredoxine.
Opm.: De fylogenetische relatie tussen Streptophyta en landplanten blijft onbekend! De Charophytina (de kranswieren) werden lange tijd beschouwd als voorlopers van de landplanten maar dit blijkt op grond van moleculaire gegevens niet te kloppen!
Embryophyta met
1. Bryophyta (mossen).
Ze leefden vanaf Devoon/Carboon.
Zijn ze ontstaan uit de voorlopers van groene algen?
2. Rhyniophytina (b.v. Cooksonia) leefden vanaf Siluur/Devoon. De stelen zijn bladerloos en dichotoom vertakt. Ze zijn isospoor. De sporophytgeneratie overheerst. Ze kwamen wereldwijd voor. De eveneens uitgestorven nauwverwante groep Zosterophyllopsida (b.v. Asteroxylon) vormen een parafyletische groep. Eveneens dichotomie en isosporie.
3. Lycopodiopsida (wolfsklauwen) zijn isospoor en de sporofyllen zijn dichotoom vertakt. Afwisselend microphylli met en zonder sporangium. Ondergrondse prothallia leven in symbiose met Fungi.
Ze bezitten floëem.
4. Selaginellales (b.v. Engels mos) komen nu nog overal voor met ongeveer 700 soorten. Bij de Selaginellales vinden we megasporangia en microsporangia. De mannelijke gametofyt blijft in de spore en de vrouwelijke gametofyt groeit uit de spore.
5. Lepidodendrales groeiden uit tot hoge bomen tijdens het Carboon. O.a. Sigillaria.
Sporangia groeien op een sporofyl die samen een strobilus vormen.
6. Isoëtales (biesvarens) zijn veelal aquatisch.
Met megasporofyllen en microsporofyllen. Isoëtales bezitten cambium.
7. Monilophyta (varens). Met 300 geslachten en 9000 soorten. Wereldwijd verspreid en vooral in schaduwrijke plaatsen. Vaak met wortelstokken Varens hebben primaire plastiden.
Het studieboek van Jens Boeningk c.s. neemt de Monilophyta samen met de Spermatophyta (zaadplanten) als twee zustergroepen.
De varens vertonen concentrische bouw van de vaatbundels met xyleen binnenin en floëem daar omheen.
8. Gymnospermen.
De voorlopers van de Gymnospermen zouden de uitgestorven Progymnospermen zijn. Zij vormden geen zaden maar bezaten wel cambium dat secundair xyleem en secundair floëem vormde.
Gymnospermen waren in eerste instantie houtachtige planten. Kruidachtige Gymnospermen zouden daaruit later zijn ontstaan.
Bij de Gymnospermen is de gametophyt sterk gereduceerd.
Tot de Gymnospermen worden gerekend:
a. Cycadopsida: de Palmvarens die zeer uitgebreid voorkwamen in het
Mesozoïcum. Nu vooral in tropische gebieden met 300 soorten. Ontstaan
tijdens Carboon. Spermatozoïden met flagel.
b. Ginkgoöpsida: Ook ontstaan in Carboon en wijd verspreid in
Mesozoïcum. Dichotome bladnervatuur. Nog steeds beschouwd als
oorspronkelijk? Levend fossiel? Spermatozoïden met flagel.
c. Cordaïtopsida: Uitgestorven. Op noordelijk halfrond verspreid
tijdens Paleozoïcum.
d. Coniferopsida: levend vanaf Carboon. Met naaldvormige of
schubvormige bladeren. Hiertoe behoren de Gnetales. De
systematische plaats hiervan is erg onduidelijk. Nauw verwant aan Pinus?
Of aan "bloemplanten"? De Gnetales omvatten drie nu levende
plantengroepen: Gnetum, Welwitschia en Ephedra.
e. Glossopteridopsida: bomen die leefden tijdens Perm/Trias.
f. Benettitopsida: leefden gedurende Trias tot Krijt.
Met "bloemachtige" kegelstructuur.
Opm.: De grote vraag is welke fylogenetische verbanden er bestaan tussen de 7 bovengenoemde groepen!. De volgorde van voorkomen in de opeenvolgende aardlagen vormt een groot probleem voor diverse theorieën van de evolutietheorie.
In H. 6.1 onder 2.3 t/m 2.6 heb ik de schimmels en paddenstoelen, mossen, varens, paardenstaarten en zaadplanten uitvoeriger behandeld.
9. Angiospermen.
De Magnoliopsida zijn Angiospermen (bedektzadigen).
Kenmerken van de Angiospermen zijn:
- bezit van zeefvaten (floëem) met begeleidende cellen
- meeldraden met 2 helmhokken
- de mannelijke gametofyt bestaat uit 3 cellen
- er vindt dubbele bevruchting plaats en daardoor vorming van triploïd
endosperm (reservevoedsel)
De Magnoliopsida bestaan vanaf vroeg-Jura.
Met 230.000 soorten vormt ze de meest diverse groep van landplanten!
Tot de Angiospermen behoren de Monocotylen en de Dicotylen.
De basale groepen en de Monocotylen hebben stuifmeelkorrels met slechts 1 opening. De Eudicotylen hebben stuifmeelkorrels met 3 openingen.
Opm.: De systematische plaats en de fylogenie van de Magnoliopsida is onduidelijk.
Over de verdere indeling van de Magnoliopsida bestaat groot verschil van inzicht.
Tot de Magnoliopsida behoren:
1. Basale Angiospermen: bloemdelen afwisselend of drievoudig
gespiraliseerd, meestal geen vruchten.
Het zijn vooral houtachtige planten. Met 8600 soorten.
- ze vormen geen monophyletische groep.
- de idioblasten bevatten etherische oliën.
- de voortplantingsorganen zijn meestal gerangschikt in een spiraal.
- de vruchtbeginsels zijn meestal niet vergroeid.
- ze bezitten primaire plastiden met chlorofyl a en b
- het aantal kernen in de embryozak varieert van 4, 8 tot 9.
O.a. de Nymphaeales (o.a. de waterlelie) worden hiertoe gerekend. Ze bezitten 4 kelkbladeren (sepalen) en wel tot 50 kroonbladeren (petalen); gerangschikt in spiralen.
2. Magnoliida: meeldraden staan in spiralen. O.a. Piperales,
Laurales en Magnoliales
3. Monocotylen: o.a. lelies, grassen, orchideeën, bolgewassen
- de zaden hebben 1 cotyl (zaadlob)
- ze hebben bijwortels
- de vaatbundels zijn collateraal gesloten
- de vaatbundels liggen verspreid
- de bladeren staan verspreid
- de bladeren zijn rolrond en/of paralelnervig.
4. Ceratophyllales: de hoornbloemfam. Op vele manieren ondergebracht
in een systeem; instaande tussen Monocotylen en Chloranthales??
Stuifmeelkorrels zonder opening. Geen trachaea-xyleem.
V.b. van convergente evolutie??
5. Eudicotylen:
- de zaden hebben 2 cotylen
- stuifmeelkorrels met 3 openingen
- ze hebben hoofd- en zijwortels
- de vaatbundels zijn collateraal-open
- de bladeren staan verspreid, tegenover elkaar, kransvormig of alternerend
- bladnervatuur: hoofdnerf met zijnerven.
Opm.: Hiermee hebben we het hele rijk van de Angiospermen gehad. De indeling in vele orden en families is deels gebaseerd op moleculaire gegevens en morfologische kenmerken. De systematische plaats van orden en vooral van families vertoont vele verschillen in criteria en zijn gebaseerd op diverse opvattingen. Er is vrijwel niets te zeggen over de periode van ontstaan van de verschillende orden en families. Noch over de fylogenetische relaties!
Kenmerken als symmetrie, wel of niet vergroeidbladig, onderstandig of bovenstandig vruchtbegingel, bladstand, nervatuur, bloemstructuren etc. variëren veelal binnen elke orde.
Indelingsbegrippen als rijken, hoofd)afdelingen, klassen, orden, families etc. zijn hiermee als achterhaald beschouwd.
4 Rhizaria.
Rhizaria is een erg diverse supergroep en zeer algemeen. De meeste taxa hebben diverse typen pseudopodia. De Retaria hebben een skelet van kalk of kiezel (Foraminiferen en Radiolariën).Ze bewogen zich voort m.b.v. pseudopodia. Ze komen voor vanaf het begin van de aardgeschiedenis. De Chlorarachniophyta hebben chloroplasten! Deze hebben ze waarschijnlijk verkregen door endocytobiose van een groenwier of een roodwier. Bewijs: tussen de membranen van de plastide bevindt zich een nucleomorf. Dit zou een overblijfsel zijn van de kern van het groenwier. De zoosporen kunnen een flagel hebben!
Bij de Cercozoa komen zowel exemplaren voor met schijnvoetjes als met flagellen. Binnen deze groep heeft Paulinella een chromatofoor die afkomstig is van een cyanobacterie.
Als je alles wilt weten over de bewijsvoering rondom deze veronderstellingen lees dan op de volgende site: The ancestor of the Paulinella chromatophore obtained a carboxysomal operon by horizontal gene transfer from a Nitrococcus-like γ-proteobacterium (nih.gov)
5 Chromalveolata (Alveolata en
Stramenopiles).
De Alveolata hebben alveoli onder de membraan. Ze hebben flagellen (b.v. de Dinophyta) of trilharen (b.v. Paramecium). Ze bezitten wel of geen bladgroenkorrels. De relaties binnen de groepen als het gaat over de herkomst van plastiden is zeer ingewikkeld. Moleculaire studies vertellen de gemeenschappelijke afkomst van alle plastiden van Rhodophyta (roodwieren). Toegegeven wordt dat endocytobiose een zeer complex proces is dat de werking van vele genen en de route van vele ingewikkelde stofwisselingsprocessen betreft. Had de voorouder van Alveolata, de Stramenopiles en de Rhizaria (de SAR-clade) nu wel of geen plastiden. Kreeg de voorouder een plastide door secundaire endocytobiose en verloren vele afstammelingen deze weer? Of ontstond secundaire endocytobiose vele malen? Of verkreeg een subgroep de plastide van een roodwier dankzij secundaire endocytobiose en gaf deze de plastide door aan andere cellijnen via tertiaire endocytobiose?
Vast staat: alle plastiden van alle groepen zijn aan elkaar verwant.
En: de Dinophyta bestaan vanaf het Cambrium.
Je ziet: alles is mogelijk en weinig is zeker!
5.1 Ciliophora
Tot de laatste behoort het ons welbekende pantoffeldiertje. Met haar macro- en micronucleus. Bovendien duizenden ciliën die in een schitterend ritme de doelgerichte voortbeweging en voeding mogelijk maken. Ze bezitten geen plastiden!
5.2 Dinophyta.
Deze groep van eukaryotische 1-celligen vertoont de meest voorkomende eigenschappen van alle eukaryota. Sommige van de Dinophyta produceren licht. Evenals vuurvliegjes en vele diepzeevissen. Lichtproductie is mogelijk doordat een eiwit luciferine door een enzym luciferase aangezet wordt om licht te geven.
Dinophyten bezitten secundaire plastiden met 3 membranen + chlorofyl a en c.
Of ze hebben tertiaire plastiden met 4 membranen + chlorofyl a en c of chlorofyl a en b.
Ze kunnen zowel autotroof als heterotroof zijn. Ze bevatten ook mitochondriën.
Ze bezitten 2 flagellen: 1 loopt via een longitudinale groeve naar buiten,
1 loopt door een equatoriale groeve. Deze heeft fijne
ciliën.
De cellen bezitten wel of geen cellulose-platen.
Er vindt zowel geslachtelijke als ongeslachtelijke voortplanting plaats.
Heel bijzonder: het DNA in de kern bevat geen histonen.
Dinophyta bestaan al sinds het Cambrium; dus al vanaf het begin van de wereldgeschiedenis!
Er zijn 4000 (!!) soorten.
Ze kunnen in symbiose leven met Radiolaria, Foraminifera, Mollusken, Cnidaria en Ciliata.
Sommige zijn parasitair.
Dinophyta kunnen over honderden vierkante km's algenbloei veroorzaken. Ze produceren daarbij giftige stoffen waaraan andere waterorganismen bezwijken.
Ze bezitten ß-caroteen en xantophyl.
ß-caroteen komt voor in groente, fruit en micro-algen. Het is een provitamine voor vit. A.
5.3 Apicomplexa.
De Apicomplexa zijn zeer bijzonder gebouwde 1-celligen en zijn obligate parasieten. Een berucht v.b. is Plasmodium, de malariaparasiet. Ze hebben 1, 2 of 3 generatiewisselingen via verschillende gastheren. Met diverse vormen van geslachtelijke voortplanting. Ze maken gebruik van een apicoplast, afkomstig van een chloroplast die omgeven is door 4 membranen. Dus ze zijn binnen gekomen door secundaire endocytobiose.
5.4 Stramenopiles.
De Stramenopiles vormen een zeer gevarieerde zustergroep van de Alveolata.
Ze is gevarieerder dan alle Metazoa (dieren) en Embryophyta (planten) samen.
Ze hebben gemeenschappelijk: een voorste flagel met holle, driedelige Mastigonema (haren op de flagellen).
Vele groepen hebben chloroplasten; vele andere niet. Of sterk gereduceerd chlorofyl. De groepen zelf zijn wel monofyletisch maar hun onderlinge relaties zijn volstrekt onduidelijk.
De groepen binnen de plastidenloze Stramenopiles verschillen enorm van elkaar: ze hebben 1 flagel, 2 flagellen of vele. Ze zijn 1-cellig of kolonievormend.
5.5 Perenosporomycetes (Oomycetes).
Ze leven veelal parasitair. 400 soorten. Ze zijn veelal 1-cellig en sommige zijn veroorzakers van ernstige plantenziekten; b.v Phytophtora infestans (aardappelziekte).
De interne classificatie blijft vele vragen oproepen.
5.6 Phaeophyceae.
Bruinwieren. 1800 soorten. Tot 60 m. lange kelp.
De voortplantingscellen met 4 flagellen: 1 voor, 1 achter, 2 lateraal. Een van de twee met 2 rijen tripartite driedelige haren.
Eén plastide per cel met 4 membranen en chlorofyl a, c1 en c2. Plus de hulppigmenten ß-caroteen, fucoxanthine, diadinoxanthine en diatoxanthine.
Met als reservevoedsel Chrysolaminarine (een polysacharide).
Er komt een haploïde en een diploïde generatie voor.
Ze hebben verschillende soorten weefsels. Vaak met luchtblazen De celwanden bestaan uit cellulose samen met zouten van ingewikkelde verbindingen die samen algininezuur (een mengsel van polyuronzuren) vormen.
5.7 Chrysophyceae.
Goudalgen. Met of zonder chlorofyl. Of sterk gereduceerde plastiden. Eén of
veelcellig. 2 flagellen. Dankzij moleculaire studies samengebracht in 1 groep.
5.8 Bacillariophyceae.
Diatomeeën verschenen, zoals al eerder vermeld, pas in de Jura. Veel te laat dus(?) Diatomeeenaarde wordt nuttig toegepast in insecticiden, tandpasta en wegmarkeringen.
De cellen bezitten chlorofyl a en c + fucoxanthine. Het zijn secundaire plastiden met 4 membranen.
Sommige gameten hebben een flagel met tripartite haren.
Diatomeeën zijn de enige algencellen met een dubbel stel chromosomen (2n) in al hun cellen, uitgezonderd de gameten.
6 Hacrobia.
Haptophyta vormt wellicht een zustergroep van de SAR-clade (Alveolata, Stramenopiles en Rhizaria).
De Cryptophyta zijn misschien meer verwant aan de Archaeoplastida.
Dus de fylogenetische positie is geheel onduidelijk. Daarom wel genoemd: 'incertae sedis eukaryoten'.
Sommige soorten hebben chlorofylkorrels, andere niet.
Sommige hebben 2 flagellen, soms als haptonema om zich vast te hechten.
6.1 Haptophyta.
De Haptophyta, met 500 soorten, kunnen enorme velden met algenbloei veroorzaken met vissterfte als gevolg. Op aarde zijn ze bijna alomtegenwoordig en vormen een groot deel van alle biomassa.
De cellen van de Haptophyta zijn bedekt met celluloseplaten. Deze zijn soms versterkt door calcium. Daardoor vele fossielen in Carboon en Krijt. Ze bezitten secundaire plastiden met 4 membranen en bevatten chlorofyl a en c + fucoxanthine en diatoxanthine. Reservevoedsel is eveneens opgeslagen als chrysolaminarine. Soms ook als olie of paramylon (net als bij de Euglenida). Paramylon is een ß-1,3 polymeer van glucose.
Sommige soorten hebben een oogvlek (vgl. Euglena).
6.2 Cryptophyta.
De Cryptophyta, met 200 soorten. Deze laatsten leven bij lagere lichtintensiteiten en dus in dieper water.
Ze hebben soms wel, soms geen bladgroenkorrels. Bezit van flagellen en de structuur van celwanden kunnen onderling verschillen. De onderlinge relaties zijn onduidelijk.
Ze bezitten Chlorofyl a en c2 + fucoxanthine en diatoxanthine afkomstig van endocytobiose van een roodwier. Zetmeelkorrels en een nucleomorf (beschouwd als restant van de roodwierkern) bevinden zich tussen de 2e en 3e membraan.
Andere groepen 1-celligen leven als endosymbionten samen met Cryptophyta. Cryptophyta hebben geen celwand. In plaats daarvan een drielagige periplast die bestaat uit 2 lagen eiwitplaten met daartussen een derde eiwitmembraan. Er is een lange flagel met 2 rijen stevige bipartite haren; de kortere flagel heeft slechts 1 rij haren. Een ander geslacht heeft slechts haren op één van de twee flagellen. Vele fossielen van de Cryptophyta bezaten geen plastiden.
De belangrijkste groeve van de cellen is van binnen bezet met explosieve organellen. Daarnaast zijn injectosomen met vergelijkbare structuur verspreid aanwezig onder de periplast. Bij prikkeling stoten ze een dubbele opgerolde draad naar buiten. Waarschijnlijk als verdediging tegen vijanden.
In de zgn. SAR-clade worden de Hacrobia samengenomen met de Chromoalveolata en de Rhizaria. Omdat ze chlorofyl c bezitten.
De systematische plaats van deze groepen is echter geheel onduidelijk. De Haptophyten zouden een zustergroep van de SAR-clade kunnen zijn of meer verwant aan de Archaeoplastida (waartoe ook de Chlorophyten en de Embryophyten behoren).
Enkele samenvattende conclusies.
1. Zonder de hypothese van het proces endocytobiose valt het hele cladistische systeem in elkaar.
Afzonderlijke processen van endocytobiose zouden door experimenten aangetoond behoren te worden. Tot nu toe heb ik daar niets over gelezen. Er bestaat een vakgroep experimentele evolutie. Wellicht bieden resultaten afkomstig van die groep nog eens boeiende resultaten hieromtrent.
De endosymbiose-hypothese veronderstelt nu dat er kernvorming plaats vond door het samengaan van een bacterie met een pro-eukaryotische cel (een Archaea). De bacterie verloor b.v. z'n celwand (Hoe is dat mogelijk?) en slokte een Archaea op. Daarna verloor de Archaea z'n celwand en plasmamembraan (hoe kan dat?). Tegelijkertijd vormde de gastheerbacterie instulpingen in de celmembraan. Eventueel werd het gastheer-genoom overgeplaatst in de Archaea en daarin werd een kern en endoplasmatisch reticulum (ER) gevormd. Aldus het gefingeerde verhaal in Microbiology door Presscott, Harley en Klein.
We leren hieruit: biologische fantasie staat voor niets!!
De nieuwste favoriet voor deze "geschiedenis" is Prochloron. Deze cellen bevatten chlorofyl a en b. Een veronderstelde voorouder van Prochlorophyten en een cyanobacterie zouden samen een chloroplast gevormd kunnen hebben.
De aldus ontstane primitieve eukaryotische cel zou samen met bacteria een mitochondrium gevormd kunnen hebben. En zo ontstonden de eerste pro-chlorophyten (primitieve groene algen).
Zowel bladgroenkorrels als mitochondriën vertonen een ingewikkelde membranenstructuur waarin zeer gecompliceerde stofwisselingsprocessen verlopen. Opname van (cyano)bacteriën geeft hiervoor geen verklaring. Die hypothese verschuift trouwens alleen maar het probleem. En dan rest nog de verklaring voor het ontstaan van een perfect georganiseerd Endoplasmatisch Reticulum (inclusief de miljoenen ribosomen) in samenhang met de kern en haar chromosomen.
2. De diverse cladistische vertakkingsschema's met de onderlinge afstanden tussen de takken en de variaties in de lengtes van die takken is onvoldoende onderbouwd. Berekeningen gebaseerd op de frequenties van nucleïnezuren en de onderlinge afstanden daartussen in het chromosoom lijken onbetrouwbaar en vaak onderling tegenstrijdig. De veronderstelde samenhang hiervan met vele stofwisselingsprocessen biedt geen oplossing voor het bewijzen van ontstaan van groepen en onderlinge verwantschappen. Zelfs de sterkste computers vermogen geen uitsluitsel daaromtrent te geven!
3. Het hele terrein van 1-cellige "algen" met of zonder chloroplasten, met of zonder skelet, met of zonder zetmeel of cellulose, met of zonder flagellen of trilharen vormen een bijna onafzienbaar geheel van veronderstelde monofyletische, polyfyletische of parafyletische groepsindeling. Afgezien van redeneringen rond endocytobiose zijn nergens overtuigende evolutielijnen tussen de hoofdgroepen of subgroepen te trekken. Alle mooie cladistische lijnen (ook op het internet, Wikipedia) vragen voorafgaand geloof en een goed vertrouwen op de "wetenschap" van het systeem.
4. Als we van waterplanten, evolutionair gezien, overgaan naar landplanten is er sprake van wensdenken. De onder de Streptophyta genoemde zgn. voorbereidingen wijzen daar op!
Naast cuticula, huidmondjes en rhizoiden moeten diverse blad-, stengel- en wortelsystemen ontstaan.
Ontwikkelingslijnen, met en zonder blijvend cambium, zijn gemakkelijk te veronderstellen. Maar hoe je van (lever)mossen varens maakt en van die laatste weer paardenstaarten en zaadplanten blijft volledig onopgelost. De noodzakelijke veranderingen zijn zo fundamenteel samenhangend dat geen enkele geleerde tot nu toe een goed onderbouwd verhaal daarover heeft weten te maken. Vooral hoe dubbele bevruchting en de vorming van reservevoedsel in de zaadlobben tesamen met de structuur binnen zaden en vruchten hebben kunnen ontstaan behoort nog altijd tot natuurraadsels of -wonderen!
5. De indeling in 6 supergroepen (domeinen) is erg onbevredigend. Als we letten op de bouw en structuur van cellen, weefsels en organen is het hoogst merkwaardig om de Holomyceta in de supergroep van Unikonta te plaatsen. Samen met alle "diergroepen" t/m de Primaten.
Even merkwaardig is de plaats van Phaeophyceae, Chrysophyceae en Bacillariophyceae samen met Ciliophora (o.a. pantoffeldiertje) en Dinophyta in de supergroep Chromalveolata.
Wellicht zou het logisch zijn om alle organismen zonder weefselstructuren binnen 1 supergroep te plaatsen. Misschien zou verdere studie van celstructuren, celwanden, reservevoedsel, en stofwisselingsprocessen kunnen leiden tot een verdere indeling van een dergelijke groep.
De Porifera (sponzen) zouden daarbij een plek op de valreep kunnen innemen.
Amoeboïde cellen en slijmcellen zouden samen in 1 super(sub)groep kunnen worden geplaatst evenals cellen met versterkte celmembranen door cellulose, kalk- of kiezelskeletten.
Het bezit van chloroplasten zou kunnen fungeren als een secundair criterium voor de indeling; evenals het bezit van chlorofyl c1 of c2.
6. Letten we op mogelijke tijden van ontstaan van de grote domeinen en hoofdgroepen dan blijkt het volgende:
- Het ontstaan van alle bacteriën en eukaryoten vanuit de levenloze materie is principieel onbewijsbaar.
- Zowel binnen de Archaeoplastida (de "planten") als de Holozoa ( de "dieren") worden gigantische evolutionaire sprongen gemaakt zoals b.v. tussen "algen en mossen", tussen "mossen en varens", tussen "varens en zaadplanten", tussen "naaktzadigen en bedektzadigen", tussen 1- en 2-zaadlobbhigen, tussen "sponzen, holtedieren en ribkwallen", tussen "trilobieten, krabben, spinnen, insecten en schorpioenen", tussen "platwormen, raderdiertjes, armpotigen, ringwormen en weekdieren", tussen "graptolieten en stekelhuidigen", enz. enz. Enig fossiel bewijs is daarbij meestal niet aanwezig.
7. Alle grote groepen "water- en landplanten" bestaan voor het Krijt.
Als we er van uitgaan dat tegen het einde van het Krijt de grootste van de 5 extinctieperiodes plaats greep en alle grote groepen organismen gedurende de periode Precambrium t/m Krijt bestonden vanaf het begin van de schepping, betekent dit dat alle grote afdelingen van het plantenrijk t/m de tweezaadlobbige zaadplanten gedurende alle aardperioden aanwezig waren.
Gedurende het Trias wordt verondersteld dat Pangea bestond. Met de onderdelen Gondwana en Siberia. Er heerste toen een gelijkmatig warm klimaat en er waren grote ondiepe zeegebieden langs de kusten. Alle groepen hoogontwikkelde planten (Gymnospermen, zaadvarens en zaadplanten) en dieren (tweekleppigen, geleedpotigen, inktvissen, (kraak)beenvissen, amfibieën, reptielen (Sauriërs) en zoogdieren) worden in die periode als fossiel gevonden. Terwijl aan het einde van het Perm enorme vulkaanuitbarstingen (en andere processen) zorgden voor het uitsterven van vele zaadplanten en (zee)dieren. Tijdens de Jura blijft het klimaat tamelijk rustig. Alleen iets koeler. Bomen krijgen jaarringen! Verondersteld wordt dat er nu enorme zoutlagen en immense voorraden aardolie en aardgas zijn ontstaan.
Dat laatste vooral dankzij enorme productie van plankton en nekton in de ondiepe zeeën.
De continenten beginnen uiteen te drijven. Als dat proces vrij snel verliep ging het gepaard met vele vormen van orogenese.
In het recent (2019) verschenen boek 'Earth History and Palaeogeography' door Torsvik en Cocks worden 48 orogenesen opgesomd in Appendix 3. Dit houdt in dat vrijwel alle nu bestaande gebergten door orogenese-processen zijn gevormd.
Met de daarbij behorende processen van aardbevingen en vulkaanuitbarstingen. Met als gevolgen enorme aardverschuivingen, tsunami's, overstromingen etc. Daardoor zullen zeer vele organismen overdekt zijn en gefossiliseerd. Een min of meer logische sortering zal daarbij zijn opgetreden. Snel zwemmende dieren (vissen, sauriërs en zoogdieren) en vliegende dieren (insecten, sauriërs, vogels en zoogdieren) zullen als laatste en allerlei kruipende organismen als eerste zijn bedekt. Maar deze sortering van fossielen zal mede zijn bepaald door plaats en tijd van de gebeurtenissen.
Het ontstaan van enorme voorraden aan steenkool, aardolie en aardgas zullen vooral met de heftigste vormen van orogenese verbonden zijn geweest.
Ook het ontstaan van de geweldige zoutlagen, wereldwijd, hebben waarschijnlijk hiermee te maken gehad.
Zie hierover uitvoeriger H. 6.5
reageren? tnbuist@gmail.com
'